VeloCycle (Lederer et al., Nat Methods 2024)
Lederer, Leonardi, Talamanca, Bobrovskiy, Herrera, Droin, Khven, Carvalho, Valente, Dominguez Mantes, Mulet Arabí, Pinello, Felix Naef & Gioele La Manno. Nature Methods 21:2271–2286 (2024-10-31). EPFL (La Manno = first author of the 2018 origin paper velocyto-2018; Naef = circadian/systems-biology) + Broad/MGH (Pinello). Companion News & Views: velocycle-nv-2024.
Summary
VeloCycle reframes RNA velocity as statistical inference on a manifold-constrained generative dynamical system: instead of fitting gene-wise kinetics independently and reconciling post hoc, it jointly learns the low-dimensional expression manifold and a velocity field constrained to lie tangent to that manifold, in one Bayesian model (Pyro / SVI). Applied to the cell cycle — a 1D periodic manifold parameterized by phase φ ∈ [0,2π] with gene expression as a Fourier series — it infers an angular speed ω(φ) for cell-cycle progression. The landmark result for this wiki: by expressing velocity in mean-half-life units (rpmh) and scaling by measured average mRNA half-lives, VeloCycle reports a cell-cycle period in hours that matches live-imaging time-lapse microscopy and EdU labeling (dHF ≈ 15.3–15.8 h; RPE1 ≈ 17.7 h) — “the first example of a direct validation of RNA velocity estimation with experimental methodologies … justifies the use of VeloCycle output in units of real (not pseudo) time.” It also delivers statistical velocity inference for the first time: credibility testing for nonzero velocity, and differential-velocity significance between conditions (erlotinib drug response, genome-wide Perturb-seq). From velocyto-2018’s author, this is the origin paper’s physical-time grounding, re-established with statistical rigor — a second route to metric time distinct from dynamo’s metabolic labeling.
Figures
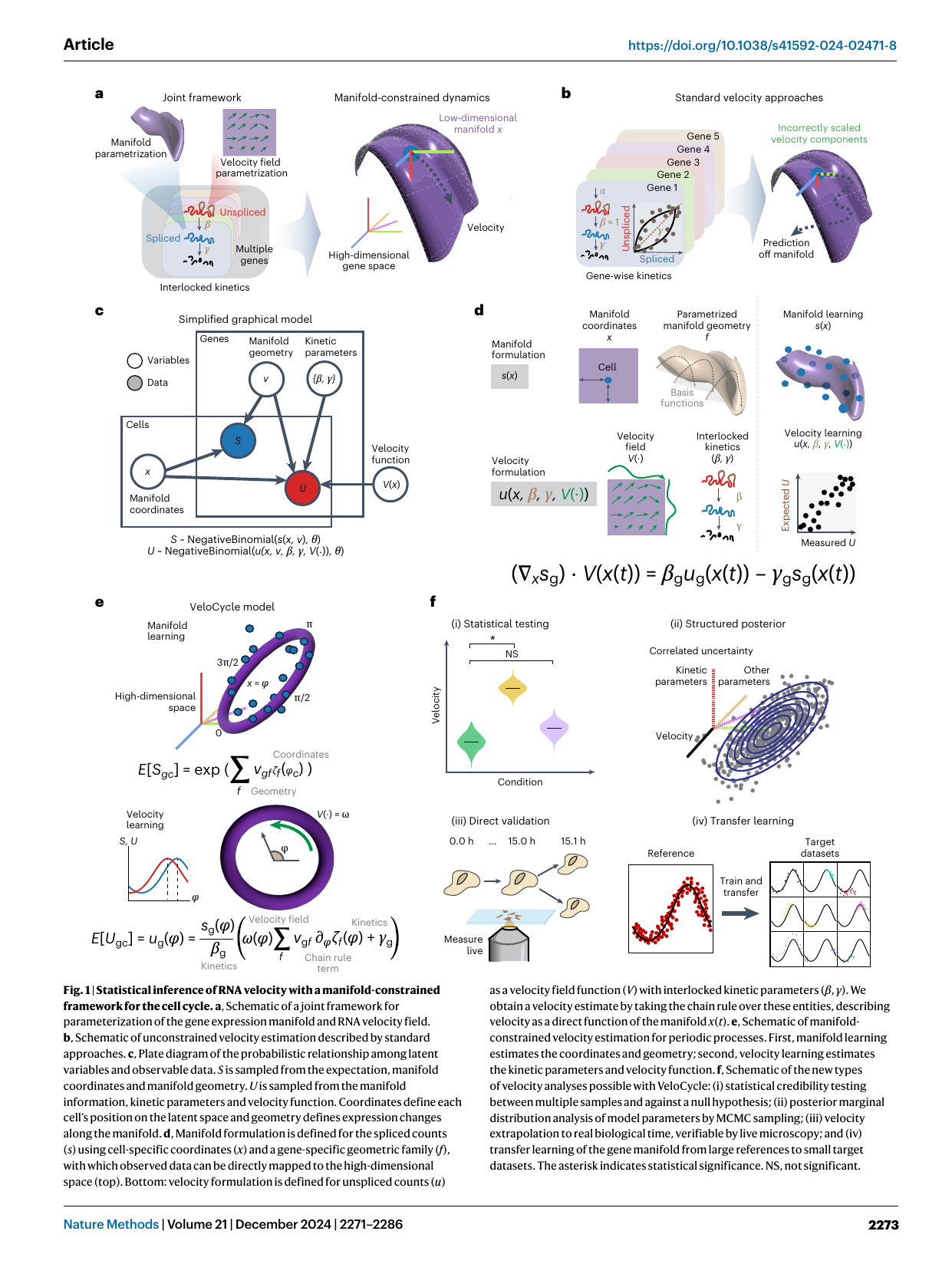
Fig 1 — manifold-constrained velocity framework (Lederer et al., Nat Methods 2024; velocycle-2024). (a) Joint estimation of the gene-expression manifold x and a velocity field V(x) constrained tangent to the manifold — vs (b) standard gene-wise velocity, whose components live on different timescales and predict off the manifold. (c) Plate diagram: spliced S and unspliced U are negative-binomial around manifold + kinetics. (e) The VeloCycle cell-cycle model: expression as a Fourier series of phase φ; velocity = angular speed ω(φ) around the ring. (f) The four new capabilities: (i) statistical velocity significance testing, (ii) structured-uncertainty posterior, (iii) direct validation against live imaging, (iv) transfer learning to small datasets. The governing identity: ∇ₓs·V(x) = β·u(x) − γ·s(x).

Fig 5 — validation in real (not pseudo) time (Lederer et al., Nat Methods 2024). VeloCycle’s velocity, scaled by measured average half-lives, yields a cell-cycle period in hours that overlaps the period measured by time-lapse microscopy (dHF 15.0 h; RPE1 16 h 40 min) and by continuous EdU labeling (RPE1 16.8 h). Panel (q) compares cell-cycle periods from VeloCycle (standard scRNA-seq), Dynamo (LAP), and Dynamo-Metabolic — VeloCycle’s posterior overlaps the gold standard without metabolic labels. This is the figure that earns “units of real time.”
Key Claims
- Manifold-constrained, dynamically consistent velocity. Velocity is parameterized as an autonomous field V(x) on manifold coordinates, forced tangent to the manifold — eliminating the gene-wise inconsistency where velocity components sit on different timescales and point off the traversed manifold. Resolves a critique GraphVelo also targets, but here by construction in a single generative model rather than as post-hoc projection.
- Cell cycle as a 1D periodic manifold. Phase φ ∈ [0,2π]; gene expression = Fourier series of φ; velocity = angular speed ω(φ) (radians per mean half-life, rpmh), allowed to vary with phase (speed modulation). Manifold learning recovers phases at circular correlation r≈0.95 vs ground truth; outperforms DeepCycle (autoencoder; 60% lower MSE).
- Metric time, validated. Scaling ω by measured average half-lives → cell-cycle period in hours, matching live-imaging and EdU ground truth across cell types with distinct periods (dHF ≈15.3–15.8 h, RPE1 ≈17.7 h). G2/M progresses faster (0.47 rpmh) than G1 (0.37) / S (0.36).
- Statistical velocity inference (first of its kind). Full Bayesian posterior (SVI; MCMC/NUTS for the correlated structure) supports (1) credibility testing for nonzero velocity, (2) differential-velocity significance between samples — demonstrated on erlotinib (slower mitotic cell cycle at D3, localized to G2/M) and genome-wide Perturb-seq knockdowns in RPE1.
- Structured uncertainty + transfer learning. A low-rank multivariate-normal posterior captures the genuine correlation between degradation γ and angular speed ω (mean-field SVI underestimates velocity uncertainty); manifold parameters transfer from large references to small/sparse target datasets.
- Spatiotemporal biology. Radial-glia cell-cycle speed varies along the developing anterior–posterior axis (FB < MB < HB); regionally defined progenitor speed differences.
Physical-time grounding (standing lens) — a rare metric-time case
- Latent time — ordinal or metric? METRIC (on the cell cycle). The phase φ is ordinal-periodic, but the inferred angular speed, scaled by measured half-lives, gives a period in hours validated against live imaging + EdU — genuine metric time, not pseudo-time. With dynamo, one of only two methods in the wiki reaching metric time, and the first with direct real-time experimental validation.
- Rate–time scale degeneracy. Broken — by a measured-rate anchor, not labeling. Velocity is reported in mean-half-life units; converting to hours requires an externally measured average mRNA half-life (≈1 h assumed/measured). So the absolute timescale is fixed by a measured degradation rate, a different anchor than metabolic-labeling’s known Δt. The degradation–splicing rate ratio is recovered near-perfectly (r≈0.99 in simulation).
- External time anchor. Yes — three, and none is metabolic labeling for the estimate itself: (i) the cell cycle’s known periodicity (a closed 1D manifold), (ii) measured average half-lives to scale rpmh→hours, (iii) live-imaging period + EdU as validation ground truth. (scEU-seq labeling is used only for cross-comparison, not to fit the velocity.)
- Constant-rate assumptions. β, γ gene-specific constants (jointly inferred), but the angular speed ω(φ) is phase-dependent — it relaxes the constant-speed assumption along the cycle (the “speed modulations” of the title), the axis most relevant to physical-time mapping.
The decisive entry for the wiki’s thesis. The “temporal axis is open / only dynamo reaches metric” framing must now be qualified: on a process with known periodicity and measured rate scale (the cell cycle), metric time is reached and experimentally validated — by velocyto-2018’s own author, closing the loop on the origin reframe (physical→ordinal→back to physical). The general (non-periodic, developmental) trajectory remains open. For FlowVelo: metric time is attainable when (a) the manifold geometry is constrained/known and (b) the rate scale is anchored by a measured rate (half-life) or labeling — VeloCycle is the proof of principle to extend beyond the cell cycle.
Key Quotes
“this is the first example of a direct validation of RNA velocity estimation with experimental methodologies and justifies the use of VeloCycle output in units of real (not pseudo) time.” — Results.
“we introduce a Bayesian model of RNA velocity that couples velocity field and manifold estimation in a reformulated, unified framework, identifying the parameters of an explicit dynamical system.” — Abstract.
Connections
- VeloCycle — the method entity.
- velocyto-2018 / GioeleLaManno — the 2018 origin and its author; VeloCycle re-grounds physical time.
- physical-time-grounding — a second route to metric time (periodic manifold + measured half-lives + live-imaging), beside dynamo’s labeling.
- metabolic-labeling — the alternative anchor it does not need (validates against EdU/scEU-seq instead).
- manifold-consistent-velocity / GraphVelo — manifold-tangent velocity, here by construction in one generative model.
- latent-time — periodic phase that becomes metric via angular speed + half-life scaling.
- veloVI / Cell2fate — the Bayesian-uncertainty lineage; VeloCycle adds significance testing.
- dynamo — the other metric-time method; VeloCycle matches its LAP period estimate without labels.
- velocity-skepticism — a constructive answer (statistical control + manifold consistency) to the “heuristics / not manifold-consistent” critiques.
- FelixNaef — co-corresponding author (cell-cycle / circadian systems biology).
- velocycle-nv-2024 — the News & Views commentary.
- FlowVelo — proof that metric time is reachable with constrained geometry + a rate anchor; extend beyond the cell cycle.
Contradictions
- Updates the wiki’s central “temporal axis is open” claim. Prior pages framed metric time as reachable only via metabolic-labeling (dynamo), with everything else ordinal. VeloCycle reaches validated metric time on the cell cycle without labeling. The corrected framing: the temporal axis is solved on periodic/known-geometry processes with a measured rate scale, still open for general developmental trajectories. Propagated to physical-time-grounding, overview, and physical-time-grounding-across-methods.