Monod (Gorin et al., Nature Methods 2025)
Gorin, Chari, Carilli, Vastola & Pachter. Nature Methods 22:2286–2300 (2025-11-07). Caltech /
Broad / Harvard. Python package Monod. GennadyGorin’s PhD centerpiece; the constructive
biophysics pole of velocity-skepticism. Full text now in raw/.
Summary
Monod fits stochastic biophysical (chemical-master-equation) models of transcription directly to the joint distribution of nascent + mature single-cell counts, explicitly separating a biological component (transcription k, splicing β, degradation γ, burst size) from a technical component (capture, sampling). By exploiting stochasticity and the nascent/mature pairing, it identifies regulation invisible to mean-expression DE, compares mechanistic hypotheses, integrates across technologies, and minimizes distortive normalization. It is not a velocity or latent-time method — it is a measurement-and-kinetics model — but it bears directly on velocity because it attacks the counts and normalization upstream of every velocity estimate, and surfaces a concrete bias: nascent capture is gene-length-dependent, which skews the very u/s ratio velocity is built on.
Key Claims
- Two-component model. Biological: ∅ →k X_N →β X_M →γ ∅ (transcription→splicing→degradation). Technical: nascent capture rate C_NL ∝ gene length L; mature capture λ_M length-independent. Standard pipelines wrongly treat nascent and mature as independent, or ignore nascent.
- Six transcription models, incl. constitutive (Poisson), bursty (geometric burst size — the recommended default), extrinsic (gamma-distributed k from cell–cell heterogeneity), Cox–Ingersoll–Ross. “One does not expect any single model to always perform best.”
- Rates in units of k. Assumes approximate steady state and fits rates in units of the transcription rate (k ≡ 1) — i.e. parameters are relative, echoing the rate-scale relativity the wiki tracks (here at the steady-state-distribution level, not the time level).
- Inference + uncertainty. Technical params shared genome-wide (grid search); biological params by gradient descent on KL divergence; uncertainty via the Fisher information matrix; goodness-of-fit by χ² / Hellinger.
- DE-θ. Generalizes differential expression to differential biophysical parameters (burst size, burst frequency, splicing, degradation), detecting noise modulation that keeps the mean constant — invisible to standard DE. Demonstrated on IdU DNA-damage, glutamatergic vs GABAergic neurons, germ-cell splicing, PDAC drug resistance, radiation-injury T cells.
Physical-time grounding (standing lens)
Monod sits upstream of and orthogonal to the four axes — it models steady-state count distributions, not velocity or time — but it constrains their inputs:
- Latent time. N/A — no time/velocity inferred; Monod returns kinetic parameters + uncertainty.
- Scale degeneracy. Echoed at the rate level: rates are fit in units of k (k≡1), so they are relative — the steady-state-distribution analog of velocity’s scale degeneracy.
- External anchor. None of its own (steady-state snapshot); but its technical model is the kind of explicit accounting velocity methods lack.
- Constant-rate / normalization. Its core contribution: the u/s counts feeding the splicing-kinetics-ode are distorted by normalization and by length-biased nascent capture — so the α/β/γ (and hence velocity and latent time) inherit the distortion.
The precise Monod takeaway for FlowVelo: before arguing about time, fix the counts. Nascent capture is length-biased and nascent/mature are causally linked — modeling that (rather than normalizing it away) is prerequisite to trustworthy α/β/γ. Borrow the biological-vs-technical split and FIM uncertainty ideas.
Figure
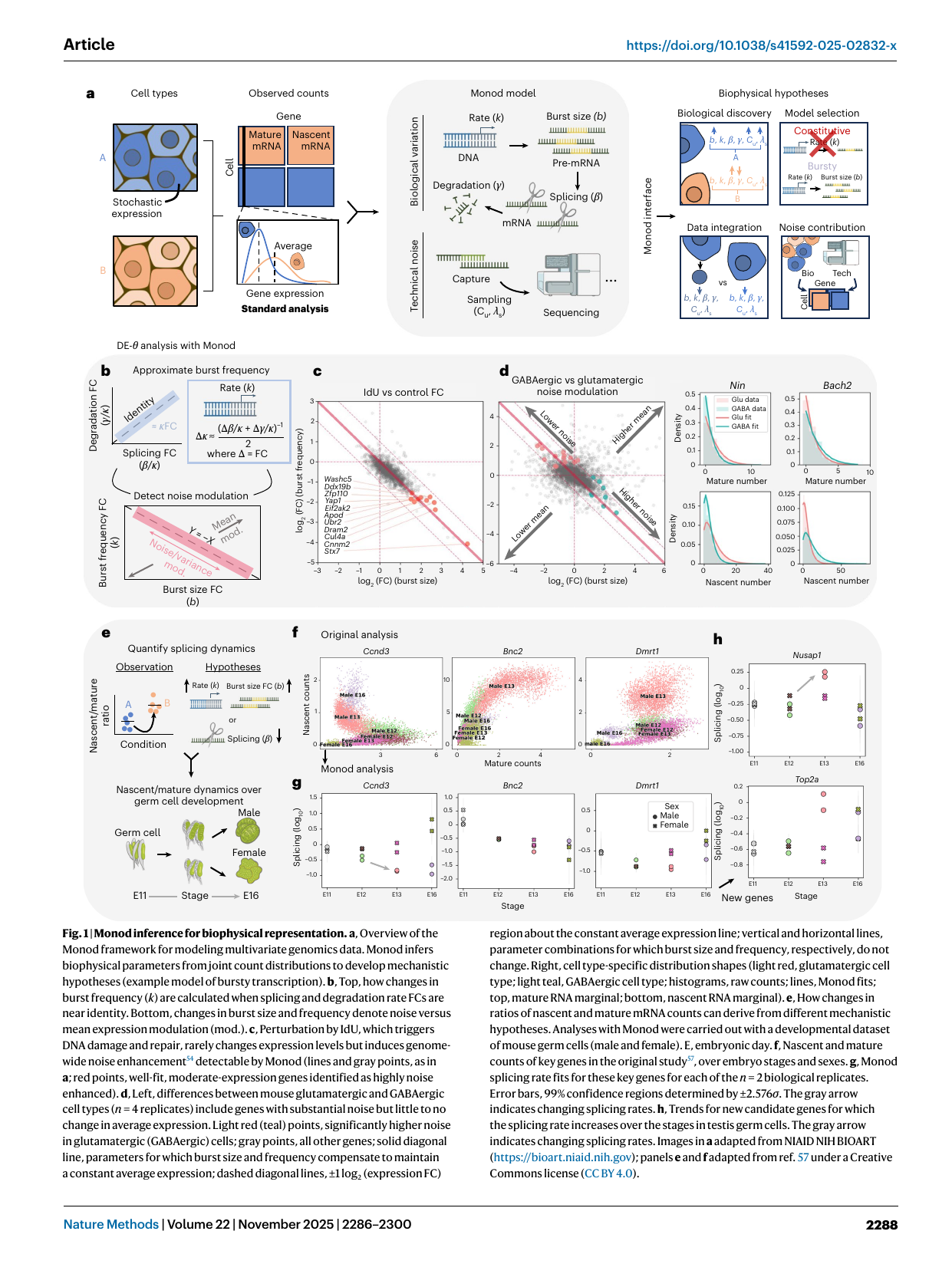
Fig 1 — the Monod framework (Gorin et al., Nat Methods 2025; monod-gorin). (a) From observed nascent + mature counts, Monod fits a model with a biological block (rate k → burst size b → DNA → splicing β → degradation γ) and a technical block (capture C_N, sampling), then exposes a “Monod interface” for biological discovery, model selection (constitutive vs bursty), data integration, and noise-contribution decomposition (bio vs technical, per gene). (b–d) DE-θ: changes in burst size vs burst frequency and noise modulation at constant mean. (e–h) splicing-rate dynamics over germ-cell development. The figure is the visual case for “model the stochasticity, don’t normalize it away.”
Key Quotes
“an alternative, physical approach to leverage the stochasticity, size and multimodal nature of these data to explicitly distinguish their biological and technical facets … minimize the use of opaque or distortive normalization and transformation techniques.” — Abstract.
“Common bioinformatic analysis pipelines often treat nascent and mature mRNA as independent rather than causally linked, conflate them or ignore nascent mRNA entirely.” — the velocity-relevant indictment.
Connections
- Cell2fate — the method entity (upgraded from bookmark-only).
- GennadyGorin / LiorPachter — authors; the biophysics pole of velocity-skepticism.
- splicing-kinetics-ode — the kinetics Monod models stochastically (CME); whose input counts it corrects.
- velocity-skepticism — the normalization/biophysics critique, made constructive.
- dynamical-systems-formulation — shares the “rigorous biophysics over ML heuristics” stance with JianhuaXing.
- cell-growth-omission-2025 — a sibling “missing physics” critique (growth vs normalization/capture).
- velocity-discourse-2025-2026 — bookmarked there.
- FlowVelo — fix the counts (capture bias, normalization) before the time axis.
Contradictions
- No conflict. Upgraded from abstract-level to full text: adds the technical capture model, six transcription models, DE-θ, and the k≡1 rate-relativity detail. Reinforces a measurement/normalization axis orthogonal to the identifiability/physical-time axis.