ddHodge
中文導讀
ddHodge(Maehara & Ohkawa,Kyushu,Nat Commun 2025,Julia package)是一個 vector-field reconstruction 框架:吃 scRNA-seq + velocity,用 Hodge decomposition 把 field 拆成 gradient(potential)、divergence-free rotational(curl)、harmonic 三塊。它的招牌產出: (1) potential landscape——用真實 embryogenesis data 驗證 development 是 gradient(Waddington) 系統;(2) divergence 當 differentiation potency / cell-state stability;(3) acceleration (second-order),這是前面方法做不到的。對 physical time:跟 GraphVelo 一樣是 geometry- preserving 的 downstream layer,potential 給的是 ordinal pseudotime(而且只有在 gradient- dominated 區才可靠),scale 從 input velocity 繼承,沒有 absolute-time anchor;但它把時間的 結構 講得更細(加了 acceleration 跟 potency 兩軸)。
What it is
A geometry-preserving vector-field reconstruction framework (not a velocity estimator). It consumes a velocity field on the data manifold and returns its Hodge components plus second-order (acceleration) and Jacobian information. Built by Maehara & Ohkawa (Kyushu University), implemented in Julia.
Mechanism
- Hodge decomposition: ω = grad α + curl·β + γ on the data manifold.
- Geometry-preserving manifold: local PCA tangent spaces stitched by a sheaf (connection) Laplacian into near-parallel coordinates (avoids UMAP-embedding distortion). See manifold-consistent-velocity.
- Outputs: potential (ordering / pseudotime), divergence (stability / potency), curl & harmonic (oscillation / cycle), Grassmann distance (gene-module change), Schur vectors (driver-gene cross-sections), and acceleration.
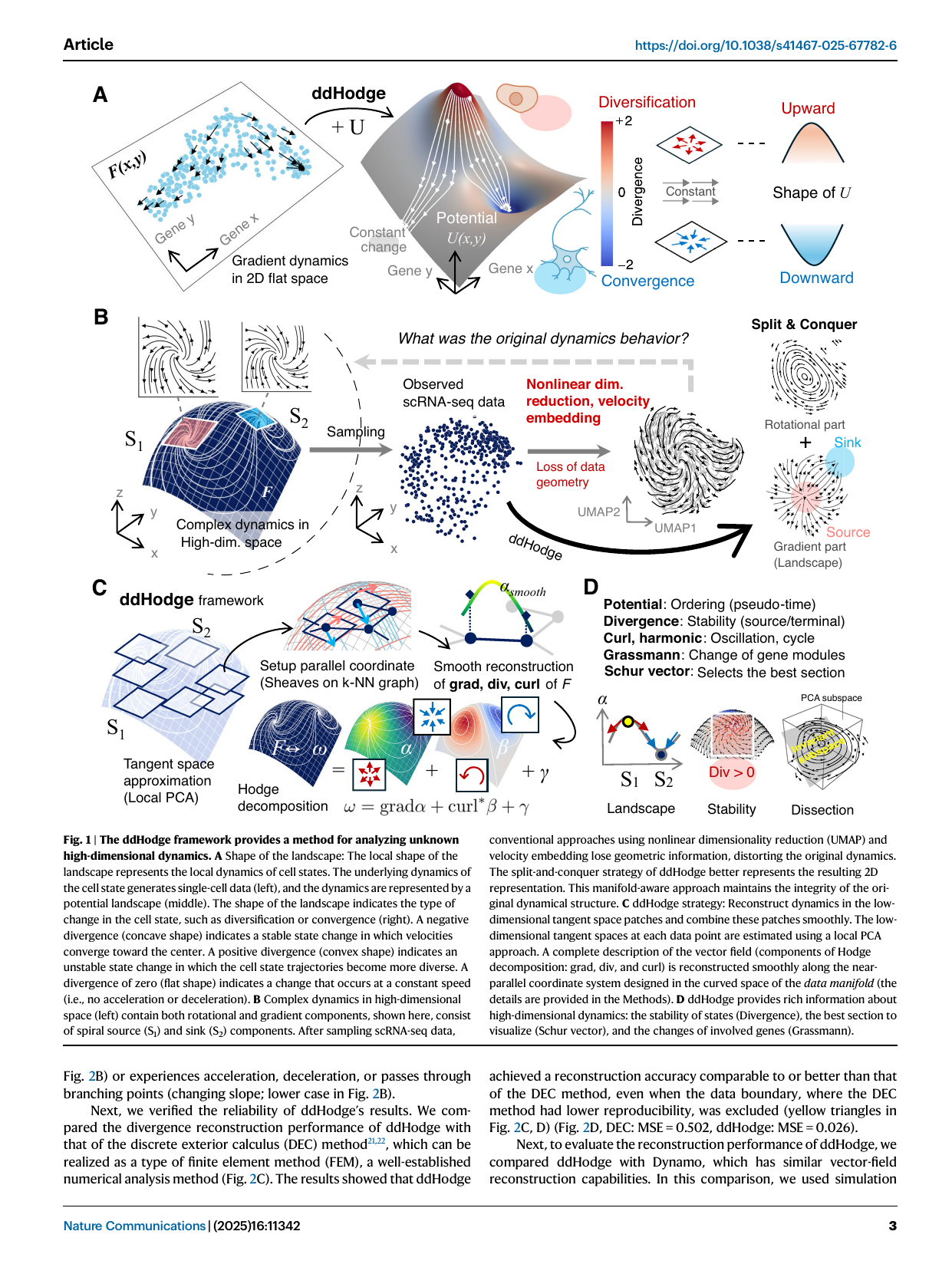
Fig 1 — ddHodge framework (Maehara & Ohkawa, Nat Commun 2025; ddhodge). Gradient dynamics define a potential landscape (A) whose local shape encodes divergence — convex/positive = diversification (branching imminent), concave/negative = convergence (canalized). “Split & conquer” (B) recovers the gradient (landscape) and rotational parts from sampled scRNA velocity, where naïve UMAP velocity-embedding would destroy the geometry. The framework (C) approximates local tangent spaces by PCA, stitches them with a sheaf (connection) Laplacian, and performs the hodge-decomposition ω = grad α + curl·β + γ. Panel D maps each indicator to biology (potential→ordering, divergence→stability, curl→oscillation, Grassmann→gene-module change, Schur→cross-section).
Dynamical indicators → biology
| Indicator | Reads out |
|---|---|
| Potential (gradient) | temporal ordering (pseudotime) — only if gradient-dominated |
| Divergence | cell-state stability / differentiation potency (neg = canalized) |
| Curl / harmonic | oscillation / cycle (e.g. cell cycle) |
| Grassmann distance | change of gene modules / dimensional change |
| Schur vectors | representative driver genes at a cross-section |
| Acceleration | change-of-change of cell state (constant vs accelerating speed) |
Physical-time scorecard
| Axis | ddHodge |
|---|---|
| Latent time | ordinal (potential = pseudotime); valid only in gradient-dominated regions; adds acceleration as richer structure |
| Rate scale | inherited from input velocity (relative) |
| External anchor | none (uses VASA-seq velocity, still snapshot) |
| Constant rates | agnostic; can detect rate-change events via divergence shifts |
| Verdict | geometry-preserving reconstruction; ordinal potential + potency/acceleration axes; scale-inheriting |
See physical-time-grounding, potential-landscape, hodge-decomposition.
Validated on
dyngen / 2D gradient & toggle-switch sims, Lorenz system, mouse embryogenesis (VASA-seq, E6.5–9.5, N≈46k), FUCCI cell cycle, muscle regeneration. Benchmarks beat DEC (FEM) and dynamo’s SparseVFC on divergence/curl/Jacobian.
Relation to other methods
- Same family as dynamo and GraphVelo (vector-field reconstruction / refinement downstream of velocity estimation), but does full Hodge decomposition + acceleration.
- Contrasts with Graph-Dynamo (Fokker–Planck transition probabilities).
- Provides the potential-landscape evidence relevant to FlowVelo’s framing.
Related
hodge-decomposition · potential-landscape · manifold-consistent-velocity · RNA velocity · splicing-kinetics-ode · dynamo · GraphVelo · latent-time · physical-time-grounding · FlowVelo